Общие сведения
Границы группы очерчены нечётко: многие грибы, способные вегетативно размножаться в одноклеточной форме и идентифицируемые поэтому как дрожжи, на других стадиях жизненного цикла образуют развитый мицелий, а в ряде случаев и макроскопические плодовые тела. Раньше такие грибы выделяли в особую группу дрожжеподобных, но сейчас их все обычно рассматривают вместе с дрожжами. Исследования 18S рРНК показали близкое родство с типичными дрожжами видов, способных к росту только в виде мицелия.
Размеры дрожжевых клеток обычно составляют 3—7 мкм в диаметре. Есть данные, что некоторые виды способны вырастать до 40 мкм[1].
Дрожжи имеют большое практическое значение, особенно пекарские или пивные дрожжи (Saccharomyces cerevisiae). Некоторые виды являются факультативными и условными патогенами. К настоящему времени полностью расшифрован геном дрожжей Saccharomyces cerevisiae (они стали первыми эукариотами, чей геном был полностью секвенирован) и Schizosaccharomyces pombe.[2]
История
Русское слово «дрожжи» имеет общий корень со словами «дрожь», «дрожать», которые применялись при описании вспенивания жидкости, зачастую сопровождающего брожение, осуществляемое дрожжами. Английское слово «yeast» (дрожжи) происходит от староанглийского «gist», «gyst», что означает «пена, кипеть, выделять газ»[3].
Дрожжи, вероятно, одни из наиболее древних «домашних организмов». Тысячи лет люди использовали их для ферментации и выпечки. Археологи нашли среди руин древнеегипетских городов жернова и пекарни, а также изображение пекарей и пивоваров. Предполагается, что пиво египтяне начали варить за 6000 лет до н. э.
, а к 1200 году до н. э. овладели технологией выпечки дрожжевого хлеба наряду с выпечкой пресного[4]. Для начала сбраживания нового субстрата люди использовали остатки старого. В результате в различных хозяйствах столетиями происходила селекция дрожжей и сформировались новые физиологические расы, не встречающиеся в природе, многие из которых даже изначально были описаны как отдельные виды. Они являются такими же продуктами человеческой деятельности, как сорта культурных растений.[5]

Луи Пастер — учёный, установивший роль дрожжей в спиртовом брожении
В 1680 году голландский натуралист Антони ван Левенгук впервые увидел дрожжи в оптический микроскоп, однако не распознал в них из-за отсутствия движения живых организмов [6]. И лишь в 1857 году французский микробиолог Луи Пастер в работе «Mémoire sur la fermentation alcoholique» доказал, что спиртовое брожение — не просто химическая реакция, как считалось ранее, а биологический процесс, производимый дрожжами[7][8].
![]()
В 1881 году Эмиль Христиан Хансен, работник лаборатории датской компании 1883 году впервые использовал её для получения пива вместо нестабильных заквасок[4]. В конце XIX века при его участии создаётся первая классификация дрожжей, в начале XX века появляются определители и коллекции дрожжевых культур.
До середины XX века учёные наблюдали только половой цикл аскомицетных дрожжей и рассматривали их всех как обособленную таксономическую группу сумчатых грибов. Японскому микологу Исао Банно в 1969 году удалось индуцировать половой цикл размножения у Rhodotorula glutinis, которая является базидиомицетом.
24 апреля1996 года года было объявлено, что Saccharomyces cerevisiae стал первым эукариотическим организмом, чей геном (12 млн пар оснований) был полностью секвенирован[10]. Секвенирование заняло 7 лет, и в нём принимали участие более 100 лабораторий[11]. Следующим дрожжевым организмом и шестым эукариотом с полностью расшифрованным геномом в 2002 году стал Schizosaccharomyces pombe[12] с 13,8 млн пар оснований.
Аскомицетные и базидиомицетные дрожжи
Различить дрожжи, принадлежащие к разным отделам грибов можно как по характеристикам их жизненного цикла, так и без его наблюдения по признакам аффинитета. К ним относится: синтез каротиноидов (встречается только у базидиомицетных дрожжей), тип убихинонов (с 5—7 изопреноидными остатками у аскомицетных и с 8—10 у базидиомицетных, хотя есть исключения), тип почкования (см.
Типичное разделение
Аскомицеты
- Saccharomycotina
- Taphrinomycotina
Базидиомицеты
- Urediniomycetes
Дрожжи способны расти на средах с низкими pH (5,5 и даже ниже), особенно в присутствии углеводов, органических кислот и других легко утилизируемых источников органического углерода[36]. Они хорошо развиваются при температурах 5—10°С, когда мицелиальные грибы уже неспособны к росту.
В процессе жизнедеятельности дрожжи метаболизируют компоненты пищевых продуктов, образуя собственные специфические конечные продукты метаболизма. При этом физические, химические и, как следствие, органолептические свойства продуктов изменяются — продукт «портится»[37]. Разрастания дрожжей на продуктах нередко видны невооруженным глазом как поверхностный налёт (например, на сыре или на мясных продуктах) или проявляют себя, запуская бродильный процесс (в соках, сиропах и даже в достаточно жидком варенье).
Дрожжи рода Zygosaccharomyces уже долгое время являются одними из важнейших агентов порчи продукции пищевой промышленности. Особенно затрудняет борьбу с ними тот факт, что они могут расти в присутствии высоких концентраций сахарозы, этанола, уксусной кислоты, бензойной кислоты и диоксида серы[38], являющихся важнейшими консервантами.
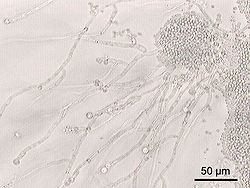
Candida albicans, образующая скопления дрожжевых клеток и псевдомицелий
Некоторые виды дрожжей являются факультативными и условными патогенами, вызвая заболевания у людей с ослабленной иммунной системой.
Дрожжи рода Candida являются компонентами нормальной микрофлоры человека, однако при общем ослаблении организма травмами, ожогами, хирургическим вмешательством, длительном применении антибиотиков, в раннем детском возрасте и в старости и т. д. грибы рода кандида могут массово развиваться, вызывая заболевание — кандидоз.
Существуют различные штаммы этого гриба, в том числе достаточно опасные. В нормальных условиях в человеческом организме дрожжи рода Candida ограничиваются в своём развитии естественной бактериальной микрофлорой человека (лактобактерии и пр.), но при развитии патологического процесса многие из них образуют высокопатогенные сообщества с бактериями.
Cryptococcus neoformans вызывает криптококкоз, особенно опасный для ВИЧ-инфицированных людей: среди них заболеваемость криптококкозом достигает 7—8 % в США и 3—6 % в Западной Европе. Клетки C. neoformans окружены прочной полисахаридной капсулой, которая препятствует их распознаванию и уничтожению лейкоцитами. Дрожжи этого вида наиболее часто обнаруживаются в помётептиц, при том что сами птицы не болеют.
Род Malassezia включает облигатных симбионтов теплокровных животных и человека, не встречающихся нигде, кроме их кожных покровов. При нарушениях иммунитета вызывают питириаз (пёстрый лишай), фолликулит и себорейный дерматит. У здоровых людей при нормальном функционировании сальных желез Malassezia никак себя не проявляют и даже играют положительную роль, препятствуя развитию более опасных патогенов.
Особенности метаболизма
Дрожжи являются хемоорганогетеротрофами и используют органические соединения как для получения энергии, так и в качестве источника углерода. Им необходим кислород для дыхания, однако при его отстутствии многие виды способны получать энергию за счёт брожения с выделением спиртов (факультативные анаэробы).
В отличие от бактерий, среди дрожжей нет облигатных анаэробов, гибнущих при наличии кислорода в среде. При пропускании воздуха через сбраживаемый субстрат дрожжи прекращают брожение и начинают дышать (поскольку этот процесс эффективнее), потребляя кислород и выделяя углекислый газ. Это ускоряет рост дрожжевых клеток (эффект Пастера). Однако даже при доступе кислорода в случае высокого содержания глюкозы в среде дрожжи начинают её сбраживать (эффект Кребтри).[13]
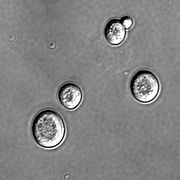
Дрожжи достаточно требовательны к условиям питания. В анаэробных условиях дрожжи могут использовать в качестве источника энергии только углеводы, причём в основном гексозы и построенные из них олигосахариды. Некоторые виды (Pichia stipitis, Pachysolen tannophilus) усваивают и пентозы, например, ксилозу.
Schwanniomyces occidentalis и Saccharomycopsis fibuliger способны сбраживать крахмал, Kluyveromyces fragilis — инулин. В аэробных условиях круг усваиваемых субстратов шире: помимо углеводов также жиры, углеводороды, ароматические и одноуглеродные соединения, спирты, органические кислоты. Гораздо больше видов способно использовать пентозы в аэробных условиях. Тем не менее, сложные соединения (лигнин, целлюлоза) для дрожжей недоступны.
Источниками азота для всех дрожжей могут быть соли аммония, примерно половина видов имеет нитратредуктазу и может усваивать нитраты. Пути усвоения мочевины различны у аскомицетовых и базидиомицетовых дрожжей. Аскомицетовые сначала карбоксилируют её, затем гидролизуют, базидиомицетовые — сразу гидролизуют уреазой.
Для практического применения важны продукты вторичного метаболизма дрожжей, выделяемые в малых количествах в среду: сивушные масла, ацетоин (ацетилметилкарбинол), диацетил, масляный альдегид, изоамиловый спирт, диметилсульфид и др. Именно от них зависят органолептические свойства полученных с помощью дрожжей продуктов.[14]
Распространение
Местообитания дрожжей связаны преимущественно с богатыми сахарами субстратами: поверхностью плодов и листьев, где они питаются прижизненными выделениями растений, нектаром цветов, раневыми соками растений, мёртвой фитомассой и т. д., однако они распространены также в почве (особенно в подстилке и органогенных горизонтах) и природных водах. Дрожжи (р.
Жизненный цикл
Отличительной особенностью дрожжей является способность к вегетативному размножению в одноклеточном состоянии. При сопоставлении с жизненными циклами грибов это выглядит как почкование спор или зиготы. Многие дрожжи также способны к реализации полового жизненного цикла (его тип зависит от аффинитета), в котором могут быть и мицелиальные стадии.

У некоторых дрожжеподобных грибов, образующих мицелий, возможен его распад на клетки (артроспоры). Это роды Endomyces, Galactomyces, Arxula, Trichosporon. У последних двух артроспоры после образования начинают почковаться. Trichosporon также образует вегетативные эндоспоры внутри клеток мицелия.
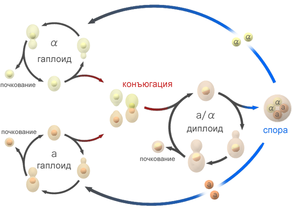
Жизненный цикл аскомицетных гапло-диплоидных дрожжей.
Наиболее характерным типом вегетативного размножения для одноклеточных аскомицетных дрожжей является почкование, лишь Schizosaccharomyces pombe размножаются не почкованием, а бинарным делением[15]. Место закладки почки является важным диагностическим признаком: полярное почкование за счёт образования шрамов почкования приводит к формированию апикулярных (лимоновидных, Saccharomycodes, Hanseniaspora, Nadsonia) и грушевидных (Schizoblastosporion) клеток;
Почкование у аскомицетных дрожжей голобластическое: клеточная стенка материнской клетки размягчается, выгибается наружу и даёт начало клеточной стенке дочерней.
Часто, особенно у аскомицетных дрожжей родов Candida и Pichia, клетки после почкования не расходятся и образуют псевдомицелий, отличающийся от истинного отчётливо видными перетяжками на месте септ и более короткими по сравнению с предшествующими конечными клетками.
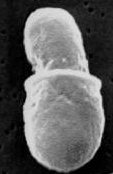
Энтеробластическое почкование у Malassezia sp.
Почкование базидиомицетных дрожжей энтеробластическое: клеточная стенка материнской клетки разрывается, из разрыва выходит почка и синтезирует свою клеточную стенку с нуля. Деление дрожжевых клеток для базидиомицетов не характерно.
Помимо обычного почкования многие виды исключительно базидиомицетных дрожжей (р. Sporidiobolus, Sporobolomyces, Bullera) способны образовывать вегетативные баллистоспоры: споры на выросте, наполненном гликогеном. Из-за гидролиза гликогена давление увеличивается и спора отстреливается на расстояние до нескольких миллиметров.

Плодовое тело Tremella mesenterica.
При половом размножении у базидиомицетов при слиянии гаплоидных дрожжевых клеток (плазмогамия) слияние ядер (кариогамия) не происходит и формируется дикариотическая клетка, дающая начало мицелию. Уже на мицелии происходит кариогамия и образуются базидиоспоры, часто даже на плодовом теле (порядок Tremallales). Единственными дрожжами среди базидиомицетов, не образующими мицелия даже при половом цикле размножения являются Xanthophyllomyces dendrorhus.

Следует отметить, что у базидиомицетовых дрожжей типы спаривания различаются обычно не одним, а большим количеством локусов. Могут сливаться только те клетки, у которых все эти локусы различны, то есть типов спаривания больше двух.
Типы спаривания

Расположение неактивных локусов HML и HMR и активного mat на хромосоме III
После слияния образуется диплоидная клетка с генотипом а/α, которой необходимо стать бесполой, чтоб больше не сливаться, и затем осуществить мейоз. Клетка этого добивается следующим образом. Ген mat а кодирует белок а1, который выполняет две функции: он подавляет считывание мРНК для белка α1 с гена mat α, поэтому фенотип α не развивается (не синтезируются α-феромоны), но он не мешает синтезу белка α2, который репрессирует а-специфичные гены, и фенотип а тоже не развивается. Во-вторых, белки а1 и α2 вместе активируют α/а-специфичные гены, которые необходимы для осуществления мейоза.
Дрожжи могут изменять свой тип спаривания с помощью рекомбинацииДНК. Это изменение у клеток происходит с частотой примерно 10-6 на клетку. Кроме локуса mat в клетке ещё имеется по копии генов mat а и mat α: соответственно HMR(Hidden MAT Right) и HML (Hidden MAT Left).[18] Но эти локусы находятся в молчащем состоянии.
Клетка заменяет работающий локус mat на копию. При этом копия снимается с того локуса, который находится в противоположенном аллельном состоянии. За этот процесс отвечает генНО. Этот ген активен только в гаплоидном состоянии. Он кодирует эндонуклеазы, которые разрезают ДНК в локусе mat. Затем экзонуклеазы убирают участок mat и на его место встает копия HMR или HML.[19]
Применение
Некоторые виды дрожжей с давних пор используются человеком при приготовлении хлеба, пива, вина, кваса и др. В сочетании с перегонкой процессы брожения лежат в основе производства крепких спиртных напитков. Полезные физиологические свойства дрожжей позволяют использовать их в биотехнологии. В настоящее время их применяют в производстве ксилита[20], ферментов, пищевых добавок, для очистки от нефтяных загрязнений.
https://www.youtube.com/watch?v=Ct7G3XjkhtY
Также дрожжи широко используются в науке в качестве модельных организмов для генетических исследований и в молекулярной биологии. Пекарские дрожжи были первыми из эукариот, у которых была полностью определена последовательность геномной ДНК. Важным направлением исследований является изучение прионов у дрожжей.
